MÚSCULO CARDÍACO EL CORAZÓN COMO BOMBA.
 A partir de este capítulo comenzamos a estudiar el corazón
y el aparato circulatorio. El corazón que se muestra en la figura está formado
en realidad por dos bombas separadas: un corazón derecho, que bombea sangre a
los pulmones, y un corazón izquierdo, que bombea sangre a los órganos
periféricos.
A partir de este capítulo comenzamos a estudiar el corazón
y el aparato circulatorio. El corazón que se muestra en la figura está formado
en realidad por dos bombas separadas: un corazón derecho, que bombea sangre a
los pulmones, y un corazón izquierdo, que bombea sangre a los órganos
periféricos.
Fisiología del músculo
cardíaco
El corazón está compuesto por tres tipos principales de
músculo cardíaco (miocardio): músculo auricular, músculo ventricular y las
fibras musculares excitadoras y conductoras especializadas.
Los tipos de músculo auricular y ventricular se contraen en
gran medida de la misma manera que el músculo esquelético, con la diferencia de
que la duración de la contracción es mucho mayor. Por el contrario, las fibras
excitadoras y conductoras especializadas se contraen solo débilmente debido a
que contiene pocas fibrillas contráctiles; en lugar de ello, muestran ritmo y
diversas velocidades de conducción, proporcionando un sistema de estimulación
cardíaca que controla el latido rítmico.
Anatomía fisiológica
del músculo cardíaco.
La figura 1-2 presenta una imagen histológica típica del
músculo cardíaco, que muestra las fibras cardíacas dispuestas en un enrejado,
con fibras que se dividen, se reúnen y se vuelven a separar. Se observa
inmediatamente en esta figura que el músculo cardíaco es estriado, de la misma
manera que el músculo esquelético típico. Además, el músculo cardíaco tiene
miofibrillas típicas que contienen filamentos de actina y de miosina casi
idénticos a los del músculo esquelético, y estos filamentos se interdigitan y
se desplazan unos a lo largo de los otros, de la misma manera que el músculo
esquelético. En otros aspectos, el músculo cardíaco es, como veremos bastante
diferente del músculo esquelético.
El músculo cardíaco
como sincitio.
Las áreas oscuras que las fibras musculares cardíacas en la
figura 1-2 se denominan discos intercalares; se trata realmente de membranas
celulares que separan entre si las células musculares cardíacas individuales.
Es decir, las fibras musculares cardíacas están compuestas por muchas células
individuales conectadas en serie entre sí. Sin embargo, la resistencia
eléctrica que ejercen los discos intercalares es tan solo 1/400


Potenciales de acción
en el músculo cardíaco
 El
potencial de acción que se registra en una fibra muscular ventricular, que se
muestra en la figura 9-3, es en promedio de aproximadamente 105 mV, lo que
significa que el potencial intracelular aumenta desde un valor muy negativo, de
aproximadamente –85 mV, entre los latidos hasta un valor ligeramente positivo,
de aproximadamente +20 mV, durante cada latido. Después de la espiga inicial la
membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una
meseta, seguida al final de la meseta de una repolarización súbita. La
presencia de esta meseta del potencial de acción ha- ce que la contracción
ventricular dure hasta 15 veces más en
el músculo cardíaco que en el músculo esquelético.
El
potencial de acción que se registra en una fibra muscular ventricular, que se
muestra en la figura 9-3, es en promedio de aproximadamente 105 mV, lo que
significa que el potencial intracelular aumenta desde un valor muy negativo, de
aproximadamente –85 mV, entre los latidos hasta un valor ligeramente positivo,
de aproximadamente +20 mV, durante cada latido. Después de la espiga inicial la
membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una
meseta, seguida al final de la meseta de una repolarización súbita. La
presencia de esta meseta del potencial de acción ha- ce que la contracción
ventricular dure hasta 15 veces más en
el músculo cardíaco que en el músculo esquelético.
Al menos dos diferencias importantes entre
las propiedades de la membrana del músculo cardíaco y esquelético son
responsables del potencial de acción prolongado y de la meseta del músculo
cardíaco.
Mientras
en el musculo esquelético el potencial de acción está producido por canales
rápidos de sodio.
En
el músculo cardíaco, el potencial de acción está producido por la apertura de
dos tipos de canales:
1) Los
mismos canales rápidos de sodio activados por el voltaje que en el músculo
esquelético.
2) Otra
población totalmente distinta de canales de calcio de tipo L (canales lentos de
calcio), que también se denominan canales de calcio-sodio. Esta segunda
población de canales difiere de los canales rápidos de sodio en que se abren
con mayor lentitud y, lo que es incluso más importante, permanecen abiertos
durante varias décimas de segundo. Durante este tiempo fluye una gran cantidad
de iones tanto calcio como sodio a través de estos canales hacia el interior de
la fibra muscular cardíaca, y esta actividad mantiene un período prolongado de
despolarización, dando lugar a la meseta del potencial de acción. Además, los
iones calcio que entran durante esta fase de meseta activan el proceso
contráctil del músculo, mientras que los iones calcio que producen la contracción
del músculo esquelético proceden del retículo sarcoplásmico intracelular.

La
segunda diferencia funcional importante entre el músculo cardíaco y el músculo
esquelético que ayuda a explicar tanto el potencial de acción prolongado como
su meseta es la siguiente: inmediatamente después del inicio del potencial de
acción, la permeabilidad de la membrana del músculo cardíaco a los iones
potasio disminuye aproximadamente cinco veces, un efecto que no aparece en el
músculo esquelético.
Fases del potencial de acción del músculo
cardíaco,
*Fase
0 (despolarización): los canales de sodio rápidos se abren cuando
la célula cardíaca es estimulada y se despolariza, el potencial de membrana se
hace más positivo. Los canales de sodio activados por el voltaje (canales de
sodio rápidos) se abren y permiten que el sodio circule rápidamente hacia el
interior de la célula y la despolarice. El potencial de membrana alcanza +20 mV
aproximadamente antes de que los canales de sodio se cierren.
Fase 1 (repolarización inicial): los
canales de sodio rápidos se cierran, los canales de sodio se cierran, la célula
empieza a repolarizarse y los iones potasio salen de la célula a través de los
canales de potasio.
Fase 2 (meseta): los
canales de calcio se abren y los canales de potasio rápidos se cierran, tiene
lugar una breve repolarización inicial y el potencial de acción alcanza una
meseta como consecuencia de:
1) una mayor permeabilidad a los iones calcio
2) la disminución de la permeabilidad a los
iones potasio.
Los canales de calcio activados por el voltaje
se abren lentamente durante las fases 1 y 0, y el calcio entra en la célula.
Después, los canales de potasio se cierran, y la combinación de una reducción
en la salida de iones potasio y un aumento de la entrada de iones calcio lleva
a que el potencial de acción alcance una meseta.
Fase 3 (repolarización rápida): los
canales de calcio se cierran y los canales de potasio lentos se abren, el
cierre de los canales iónicos de calcio y el aumento de la permeabilidad a los
iones potasio, que permiten que los iones potasio salgan rápidamente de la
célula, pone fin a la meseta y devuelve el potencial de membrana de la célula a
su nivel de reposo.
Fase 4 (potencial de membrana
de reposo) con valor medio aproximado de −90 mV.
Velocidad
de la conducción de las señales en el músculo cardíaco
La
velocidad de la conducción de la señal del potencial de acción excitador a lo
largo de las fibras musculares auriculares y ventriculares es de unos 0,3 a 0,5
m/s, o aproximadamente 1/250 de la velocidad en las fibras nerviosas grandes y
en torno a 1/10 de la velocidad en las fibras musculares esqueléticas, la
velocidad de conducción en el sistema especializado de conducción del corazón,
en las fibras de Purkinje, es de hasta 4 m/s en la mayoría de las partes del
sistema, lo que permite una conducción razonablemente rápida de la señal
excitadora hacia las diferentes partes del corazón.
Período refractario del músculo cardíaco
El
músculo cardíaco, al igual que todos los tejidos excitables, es refractario a
la reestimulación durante el potencial de acción, por lo tanto el período
refractario del corazón es el intervalo de tiempo, como se muestra en la parte
izquierda de la figura 9-5, durante el cual un impulso cardíaco normal no puede
reexcitar una zona ya excitada de músculo cardíaco.
El
período refractario normal del ventrículo es de 0,25 a 0,30 s, que es
aproximadamente la duración del potencial de acción en meseta prolongado. Hay
un período refractario relativo adicional de aproximadamente 0,05 s, durante el
cual es más difícil de lo normal excitar el músculo pero, sin embargo, se puede
excitar con una señal excitadora muy intensa, como se demuestra por la
contracción «prematura» temprana del segundo . El período refractario del
músculo auricular es mucho más corto que el de los ventrículos (aproximadamente
0,15 s para las aurículas, en comparación con 0,25 a 0,30 s para los
ventrículos).
Acoplamiento
excitación-Contracción
Se refiere al mecanismo mediante el cual
el potencial de acción hace que las miofibrillas del músculo se contraigan. Al igual que en el
músculo esquelético, cuando un potencial de acción pasa sobre la membrana del
músculo cardíaco el potencial de acción se propaga hacia el interior de la
fibra muscular cardíaca a lo largo de las membranas de los túbulos transversos
(T). Los potenciales de acción de los túbulos T, a su vez, actúan sobre las
membranas de los túbulos sarcoplásmicos longitudinales para producir la
liberación de iones calcio hacia el sarcoplasma muscular desde el retículo
sarcoplásmico. En algunas milésimas de segundo más estos iones calcio difunden
hacia las miofibrillas y catalizan las reacciones químicas que favorecen el
deslizamiento de los filamentos de actina y de miosina entre sí, lo que da
lugar a la contracción muscular.
Hasta
ahora este mecanismo de acoplamiento excitación-contracción es el mismo que el
del músculo esquelético, aunque hay un segundo efecto que es bastante
diferente. Además de los iones calcio que se liberan hacia el sarcoplasma desde
las cisternas del retículo sarcoplásmico, también difunde una gran cantidad de
iones calcio adicional hacia el sarcoplasma desde los propios túbulos T en el
momento del potencial de acción, que abre los canales de calcio dependientes
del voltaje a la membrana del túbulo T. El calcio que entra en la célula activa
después los canales de liberación de calcio, también denominados canales de
receptor de rianodina, en la membrana del retículo sarcoplásmico, para activar
la liberación de calcio en el sarcoplasma.
 La fuerza de la contracción del músculo
cardíaco depende en gran medida de la concentración de iones calcio en los
líquidos extracelulares. De hecho, un corazón situado en una solución sin
calcio dejará rápidamente de latir. La razón de esta respuesta es que las
aberturas de los túbulos T atraviesan directamente la membrana de la célula
muscular cardíaca hacia los espacios extracelulares que rodean las células, lo
que permite que el mismo líquido extracelular que está en el intersticio del
músculo cardíaco se introduzca en los túbulos T. En consecuencia, la cantidad
de iones calcio en el sistema de los túbulos T (es decir, la disponibilidad de
iones calcio para producir la contracción del músculo cardíaco) depende en gran
medida de la concentración de iones calcio en el líquido extracelular. En
cambio, la fuerza de la contracción del músculo esquelético apenas se ve
afectada por cambios moderados de la concentración de calcio en el líquido
extracelular porque la contracción del músculo esquelético está producida casi
por completo por los iones calcio que son liberados por el retículo
sarcoplásmico del interior de la propia fibra muscular esquelética.
La fuerza de la contracción del músculo
cardíaco depende en gran medida de la concentración de iones calcio en los
líquidos extracelulares. De hecho, un corazón situado en una solución sin
calcio dejará rápidamente de latir. La razón de esta respuesta es que las
aberturas de los túbulos T atraviesan directamente la membrana de la célula
muscular cardíaca hacia los espacios extracelulares que rodean las células, lo
que permite que el mismo líquido extracelular que está en el intersticio del
músculo cardíaco se introduzca en los túbulos T. En consecuencia, la cantidad
de iones calcio en el sistema de los túbulos T (es decir, la disponibilidad de
iones calcio para producir la contracción del músculo cardíaco) depende en gran
medida de la concentración de iones calcio en el líquido extracelular. En
cambio, la fuerza de la contracción del músculo esquelético apenas se ve
afectada por cambios moderados de la concentración de calcio en el líquido
extracelular porque la contracción del músculo esquelético está producida casi
por completo por los iones calcio que son liberados por el retículo
sarcoplásmico del interior de la propia fibra muscular esquelética.
Los
fenómenos cardíacos que se producen desde el comienzo de un latido cardíaco
hasta el comienzo del siguiente se denominan ciclo cardíaco.
Cuando
hablamos de ciclo cardiaco podemos decir que es la secuencia de eventos
eléctricos, mecánicos, sonoros y de presión relacionados con el flujo de sangre
a través de las cavidades cardíacas.
Cada
ciclo es iniciado por la generación espontánea de un potencial de acción en el
nódulo sinusal, este nódulo está
localizado en la pared lateral de la aurícula derecha, cerca del orificio de la
vena cava superior y el potencial de acción viaja rápidamente a través de ambas
aurículas y ahí a través del fascículo
auroventricular a los ventrículos, permitiendo que las aurículas se contraigan
antes de la contracción ventricular bombeando de esta manera sangre hacia los
ventrículos antes de que comience la intensa contracción ventricular.
Por lo
tanto, las aurículas actúan como bombas de cebado para los ventrículos, y los
ventrículos a su vez proporcionan la principal fuente de potencia para mover la
sangre a través del sistema vascular del cuerpo.
Cada
ciclo cardiaco consta de 2 fases principales, un periodo de relajación que se
denomina diástole, seguido de un período de contracción denominado sístole.
Diástole o fase de relajación:
Ocurre
cuando las paredes musculares del corazón se relajan y se llenan de sangre, la
sangre tras entregar el oxígeno a todo el cuerpo retorna al corazón y entra a
ambas aurículas, a la aurícula derecha por la vena cava superior y a la
aurícula izquierda por las venas pulmonares
Cuando
el corazón está lleno nuestro marcapasos natural llamado nodo sinoauricular
lanza impulsos que viajan por las paredes musculares y estimulan la contracción
del corazón.
Sístole o fase de Contracción:
Es la
fase más activa y potente del latido en ella los impulsos enviados por el nodo
hacen que las gruesas paredes de los ventrículos se contraigan, esto aumenta la
presión en ellos y hacen que se abran las válvulas aorticas y pulmonar y la
fuerza a la sangre para entrar en las grandes arterias
Ya que
las válvulas aurículaventriculares permanecen cerradas, las paredes de los
ventrículos se relajan y el corazón está listo para comenzar un nuevo latido.

El aumento de la
frecuencia cardíaca reduce la duración del ciclo cardíaco
Cuando
aumenta la frecuencia cardíaca, la duración de cada ciclo cardíaco disminuye
incluidas las fases de contracción y relajación. La duración del potencial de
acción y el período de contracción (sístole) también decrece, aunque no en un
porcentaje tan elevado como en la fase de relajación (diástole).
Para
una frecuencia cardíaca normal de 72 latidos/min, la sístole comprende
aproximadamente 0,4 del ciclo cardíaco completo. Para una frecuencia cardíaca
triple de lo normal, la sístole supone aproximadamente 0,65 del ciclo cardíaco
completo. Esto significa que el corazón que late a una frecuencia muy rápida no
permanece relajado el tiempo suficiente para permitir un llenado completo de
las cámaras cardíacas antes de la siguiente contracción.
Relación del electrocardiograma con el
ciclo cardíaco
En
este electrocardiograma se muestran las ondas P, Q, R, S y T, que son los
voltajes eléctricos que genera el corazón, y son registrados mediante el
electrocardiógrafo desde la superficie del cuerpo.
La
onda P está producida por la propagación de la despolarización en las
aurículas, y es seguida por la contracción auricular, que produce una ligera
elevación de la curva de presión auricular inmediatamente después de la onda P
electrocardiográfica.
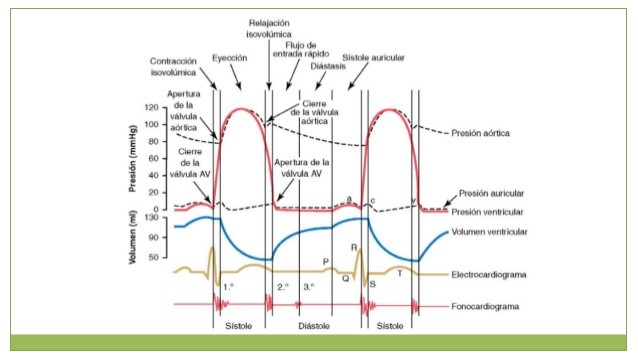 Aproximadamente
0,16 s después del inicio de la onda P, las ondas QRS aparecen como
consecuencia de la despolarización eléctrica de los ventrículos, que inicia la
contracción de los ventrículos y hace que comience a elevarse la presión
ventricular.
Aproximadamente
0,16 s después del inicio de la onda P, las ondas QRS aparecen como
consecuencia de la despolarización eléctrica de los ventrículos, que inicia la
contracción de los ventrículos y hace que comience a elevarse la presión
ventricular.
Por
tanto, el complejo QRS comienza un poco antes del inicio de la sístole
ventricular.
Finalmente,
la onda T ventricular representa la fase de repolarización de los ventrículos,
cuando las fibras del músculo ventricular comienzan a relajarse.
Por
tanto, la onda T se produce un poco antes del final de la contracción
ventricular.
Función de las aurículas como bombas de cebado para los
ventrículos
La sangre normalmente fluye de forma continua
desde las grandes ventas hacia las aurículas; aproximadamente el 80% de la
sangre fluye directamente a través de las aurículas hacia los ventrículos
incluso antes de que se contraigan las aurículas. Después, la contracción
auricular habitualmente produce un llenado de un 20% adicional de los
ventrículos. Por tanto, las aurículas actúan como bombas de cebado que aumentan
la eficacia del bombeo ventricular hasta un 20%. Sin embargo, el corazón puede
seguir funcionando en la mayor parte de las condiciones incluso sin esta
eficacia de un 20% adicional porque normalmente tiene la capacidad de bombear
entre el 300 y el 400% más de sangre de la que necesita el cuerpo en reposo.
Por tanto, cuando las aurículas dejan de funcionar es poco probable que se
observe esta diferencia salvo que la persona haga un esfuerzo; en este caso de
manera ocasional aparecen síntomas agudos de insuficiencia cardíaca,
especialmente disnea.
Función de los ventrículos como bombas
Los ventrículos se llenan de sangre durante la diástole
Durante la sístole ventricular se acumulan grandes cantidades de sangre en las
aurículas derecha e izquierda porque las válvulas AV están cerradas. Por tanto,
tan pronto como ha finalizado la sístole y las presiones ventriculares
disminuyen de nuevo a sus valores diastólicos bajos, el aumento moderado de
presión que se ha generado en las aurículas durante la sístole ventricular
inmediatamente abre las válvulas AV y permite que la sangre fluya rápidamente
hacia los ventrículos.

El período de llenado rápido dura
aproximadamente el primer tercio de la diástole. Durante el tercio medio de la
diástole normalmente solo fluye una pequeña cantidad de sangre hacia los
ventrículos; esta es la sangre que continúa drenando hacia las aurículas desde
las venas y que pasa a través de las aurículas directamente hacia los
ventrículos. Durante el último tercio de la diástole las aurículas se contraen
y aportan un impulso adicional al flujo de entrada de sangre hacia los
ventrículos. Este mecanismo es responsable de aproximadamente el 20% del
llenado de los ventrículos durante cada ciclo cardíaco.
¿Cómo funcionan las válvulas del corazón?
A la vez que el músculo del corazón se contrae
y se relaja, las válvulas se abren y se cierran, permitiendo, alternativamente,
que el flujo sanguíneo entre en los ventrículos y en los atrios. A
continuación, explicamos paso a paso cómo funcionan normalmente las válvulas
del ventrículo izquierdo:
·
Tras la contracción del
ventrículo izquierdo, la válvula aórtica se cierra y la válvula mitral se abre,
para permitir que la sangre pase desde el atrio izquierdo hasta el ventrículo izquierdo.
- Cuando se contrae el atrio izquierdo, pasa
más sangre al ventrículo izquierdo.
- Cuando el ventrículo izquierdo se contrae, la válvula mitral se cierra y la válvula aórtica se abre, para que la sangre fluya hacia la aorta.
Curva
de presión aórtica
 Cuando
el ventrículo izquierdo se contrae, la presión ventricular aumenta rápidamente
hasta que se abre la válvula aórtica. Posteriormente, después de que se haya
abierto la válvula, la presión del ventrículo aumenta mucho menos rápidamente,
porque la sangre sale inmediatamente del ventrículo hacia la aorta y después
hacia las arterias de distribución sistémica. La entrada de sangre en las
arterias hace que las paredes de las mismas se distiendan y que la presión
aumente hasta aproximadamente 120 mmHg. Después, al final de la sístole,
después de que el ventrículo izquierdo haya dejado de impulsar sangre y se haya
cerrado la válvula aórtica, las paredes elásticas de las arterias mantienen una
presión elevada en las arterias, incluso durante la diástole.
Cuando
el ventrículo izquierdo se contrae, la presión ventricular aumenta rápidamente
hasta que se abre la válvula aórtica. Posteriormente, después de que se haya
abierto la válvula, la presión del ventrículo aumenta mucho menos rápidamente,
porque la sangre sale inmediatamente del ventrículo hacia la aorta y después
hacia las arterias de distribución sistémica. La entrada de sangre en las
arterias hace que las paredes de las mismas se distiendan y que la presión
aumente hasta aproximadamente 120 mmHg. Después, al final de la sístole,
después de que el ventrículo izquierdo haya dejado de impulsar sangre y se haya
cerrado la válvula aórtica, las paredes elásticas de las arterias mantienen una
presión elevada en las arterias, incluso durante la diástole.
Relación de los tonos cardíacos con el
bombeo cardíaco
Cuando
se ausculta el corazón con un estetoscopio no se oye la apertura
de las
válvulas porque este es un proceso relativamente lento que normalmente no hace
ruido. Sin embargo, cuando las válvulas se cierran los velos de las válvulas y
los líquidos circundantes vibran bajo la influencia de los cambios súbitos de
presión, generando un sonido que viaja en todas las direcciones a través del
tórax.
Cuando
se contraen los ventrículos primero se oye un ruido que está producido por el
cierre de las válvulas AV, la vibración tiene un tono bajo y es relativamente
prolongada y se conoce como el primer tono cardíaco, cuando se cierran las
válvulas aórtica y pulmonar al final de la sístole se oye un golpe seco y
rápido porque estas válvulas se cierran rápidamente y los líquidos circundantes
vibran durante un período corto, este sonido se denomina segundo tono cardíaco.
Generación
de trabajo del corazón
 El
trabajo sistólico del corazón es la cantidad de energía que el corazón
convierte en trabajo durante cada latido cardíaco mientras bombea sangre hacia
las arterias. El trabajo minuto es la cantidad total de energía que se
convierte en trabajo en 1 min; este parámetro es igual al trabajo sistólico
multiplicado por la frecuencia cardíaca por minuto. El trabajo del corazón se
utiliza de dos maneras. Primero, la mayor proporción se utiliza, con mucho,
para mover la sangre desde las venas de baja presión hacia las arterias de alta
presión. Esto se denomina trabajo volumen-presión o trabajo externo. Segundo,
una pequeña proporción de la energía se utiliza para acelerar la sangre hasta
su velocidad de eyección a través de las válvulas aórtica y pulmonar, que es el
componente de energía cinética del flujo sanguíneo del trabajo cardíaco. El
trabajo externo del ventrículo derecho es normalmente de alrededor de la sexta
parte del trabajo del ventrículo izquierdo debido a la diferencia de seis veces
de las presiones sistólicas que bombean los dos ventrículos. El trabajo adicional
de cada uno de los ventrículos necesario para generar la energía cinética del
flujo sanguíneo es proporcional a la masa de sangre que se expulsa multiplicada
por el cuadrado de la velocidad de eyección.
El
trabajo sistólico del corazón es la cantidad de energía que el corazón
convierte en trabajo durante cada latido cardíaco mientras bombea sangre hacia
las arterias. El trabajo minuto es la cantidad total de energía que se
convierte en trabajo en 1 min; este parámetro es igual al trabajo sistólico
multiplicado por la frecuencia cardíaca por minuto. El trabajo del corazón se
utiliza de dos maneras. Primero, la mayor proporción se utiliza, con mucho,
para mover la sangre desde las venas de baja presión hacia las arterias de alta
presión. Esto se denomina trabajo volumen-presión o trabajo externo. Segundo,
una pequeña proporción de la energía se utiliza para acelerar la sangre hasta
su velocidad de eyección a través de las válvulas aórtica y pulmonar, que es el
componente de energía cinética del flujo sanguíneo del trabajo cardíaco. El
trabajo externo del ventrículo derecho es normalmente de alrededor de la sexta
parte del trabajo del ventrículo izquierdo debido a la diferencia de seis veces
de las presiones sistólicas que bombean los dos ventrículos. El trabajo adicional
de cada uno de los ventrículos necesario para generar la energía cinética del
flujo sanguíneo es proporcional a la masa de sangre que se expulsa multiplicada
por el cuadrado de la velocidad de eyección.
Habitualmente
el trabajo del ventrículo izquierdo necesario para crear la energía cinética
del flujo sanguíneo es de solo un 1% del trabajo total del ventrículo y, por
tanto, se ignora en el cálculo del trabajo sistólico total. Sin embargo, en
ciertas situaciones anormales, como en la estenosis aórtica, en la que la
sangre fluye con una gran velocidad a través de la válvula estenosada, puede
ser necesario más del 50% del trabajo
total para generar la energía cinética del flujo sanguíneo.
El
musculo cardíaco utiliza energía química para hacer el trabajo de contracción
esta energía deriva principalmente del metabolismo oxidativo de los ácidos
grasos, y en menor medida de otros nutrientes especialmente lactato y glucosa;
Por
ejemplo (la energía es necesaria para la actividad muscular) todos los
alimentos energéticos hidratos de
carbono, grasa y proteínas pueden ser oxidados en la célula y en este proceso
se liberan grandes cantidades de energía. La cantidad de energía por la
oxidación completa de un alimento, se llama "ENERGÍA LIBERADA" por
tanto la tasa de consumo de oxigeno por el corazón es una medida excelente de
la energía química liberada.
Regulación del bombeo
Cuando
una persona está en reposo su corazón bombea de 4 a 6 L por minutos, durante el
ejercicio intenso se puede exigir que bombee de cuatro a siete veces esa
cantidad. Ejemplo: el sistema nervioso durante el ejercicio emite otras señales
para ele

Regulación intrínseca
del bombeo Cardiaco: Mecanismo de Frank Starling
La cantidad de sangre bombeada por el corazón cada minuto
está determinada casi en su totalidad por el flujo de sangre procedente de las
venas al corazón, que se denomina retorno venoso. Es decir, cada tejido
periférico del organismo controla su propio flujo sanguíneo, y el total de
todos los flujos sanguíneos locales a través de todos los tejidos periféricos
que regresan por las venas a la aurícula derecha. El corazón a su vez, bombea
automáticamente a las arterias sistemáticas esta sangre que llega, de forma que
puede volver a fluir por el circuito. Esta capacidad intrínseca del corazón de
adaptarse a volúmenes de sangre que fluyen se denominan mecanismo cardiaco de
Frank Starling en honor a dos grandes fisiólogos de hace un siglo, básicamente
este mecanismo significa que cuando más se distiende el musculo cardiaco
durante el llenado mayor es la fuerza de contracción y mayor la cantidad de
sangre bombeada a la aorta.
Otra forma de expresarlo es: dentro de los limites
fisiológicos, el corazón bombea toda la sangre sin permitir que se remance una cantidad excesiva en las venas.
Explicación del
mecanismo de Frank Starling
Cuando llega a los ventrículos una cantidad adicional de
sangre, el propio musculo cardiaco se distiende
a una longitud mayor. Esto hace que el musculo se contraiga con mayor
fuerza debido a que los filamentos de actina y miosina son llevados a un grado
casi óptimo de interdigitacion para generar la fuerza, por tanto el ventrículo
a causa del aumento de bombeo propulsa de forma automática la sangre adicional
a las arterias. Está capacidad del musculo de distenderse hasta esa longitud y
de contraerse con más fuerzas es característica de todo el musculo estriado.
Existe otro factor que aumenta el bombeo cardio cuando su
volumen se incrementa la instancia de pared auricular derecha aumenta
directamente la frecuencia cardiaca de un 10 a un 20% .También esto contribuye
a aumentar la cantidad del bombeo de sangre por minuto, aunque su contribución
es mucho menor a la del mecanismo de Frank Starling.
Efecto de los iones
potasio y calcio sobre la función cardíaca.
Al estudiar los potenciales de membrana, se señaló que los
iones potasio tienen un llamativo efecto sobre los potenciales de membrana y
los potenciales de acción, y que los que los iones calcio desempeñan un papel
especialmente importante en la activación del proceso contráctil muscular. Por
tanto, cabe esperar que las concentraciones de estos dos iones en el líquido
extracelular tengan también importantes efectos sobe el bombeo cardíaco.
Efecto de los iones
potasio
El exceso de potasio en los líquidos extracelulares hace
que el corazón se dilate y quede flácido, y que también disminuya la frecuencia
cardíaca. Las cantidades grandes pueden también bloquear la conducción del
impulso cardíaco de las aurículas a los ventrículos a través del haz A-V. La
elevación de la concentración de potasio a tan solo 8 a 12 mEq/L (de dos a tres
veces el valor normal), puede causar esta debilidad del corazón y un ritmo
anormal tales que pueden causar la muerte.
Estos efectos son producidos en parte porque si
existe una concentración elevada de potasio en los líquidos extracelulares,
disminuye el potencial de membrana de reposo de las fibras musculares
cardíacas. A medida que disminuye el potencial de membrana, decrece también la
intensidad del potencial de acción, lo que debilita progresivamente la
contracción cardíaca.
Efecto de los iones de
calcio
El
exceso de iones calcio produce efectos casi exactamente opuestos a los de los
iones de potasio, haciendo que el corazón caiga en una contracción espástica.
Esto se debe al efecto directo de los iones calcio de excitación del proceso
contráctil cardíaco. A la inversa, el déficit de iones calcio causa flacidez
cardíaca, similar al efecto del potasio elevado. Afortunadamente, los niveles
sanguíneos de calcio iónico, están normalmente regulados en una banda muy
estrecha, por lo que estos efectos cardíacos de las concentraciones anormales
de calcio rara vez constituyen un motivo de preocupación en la clínica.
Efecto de la
temperatura sobre la función cardíaca
El aumento de la temperatura, como el que ocurre cuando
existe fiebre, causa un gran incremento de la frecuencia cardíaca, a veces
hasta el doble de la normal. La disminución de la temperatura produce grandes
descensos de la frecuencia cardíaca, cayendo a tan solo unos pocos latidos por
minuto cuando la persona se encuentra cerca de la muerte por hipotermia, a
temperaturas de 15.5 a 21.1 °C (60 a 70
°F). Presumiblemente estos efectos son consecuencia de que el calor aumenta la
permeabilidad de la membrana muscular cardíaca a los iones controladores, lo
cual tiene como consecuencia una aceleración del proceso de autoexcitación.
La fuerza contráctil del corazón con frecuencia
resulta temporalmente potenciada por un aumento moderado de la temperatura,
pero una elevación prolongada de la temperatura agota los sistemas metabólicos
del corazón y causa debilidad. Por ello la función óptima del corazón depende
en gran medida de un control adecuado de la temperatura corporal por los
mecanismos termorreguladores.


